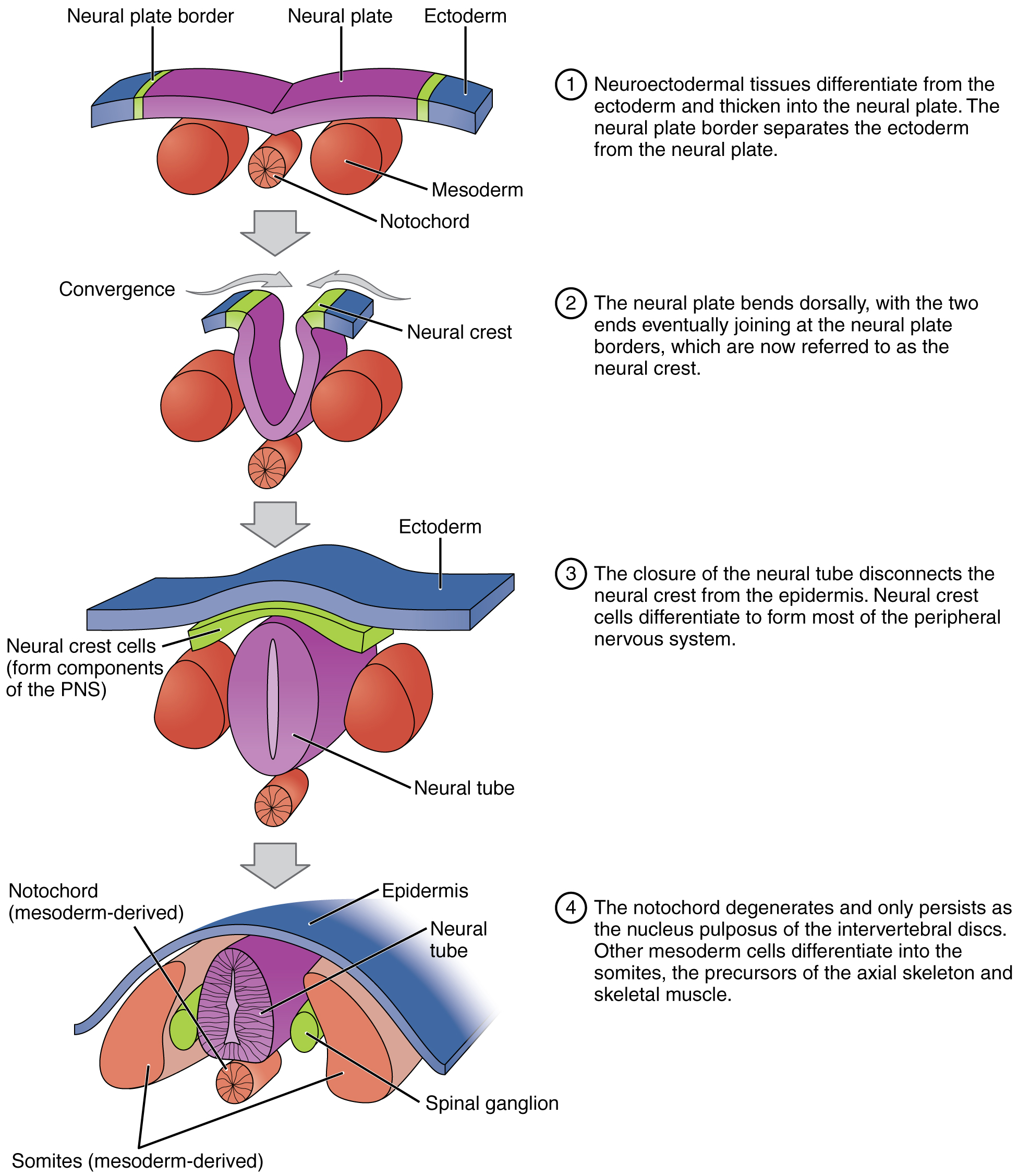
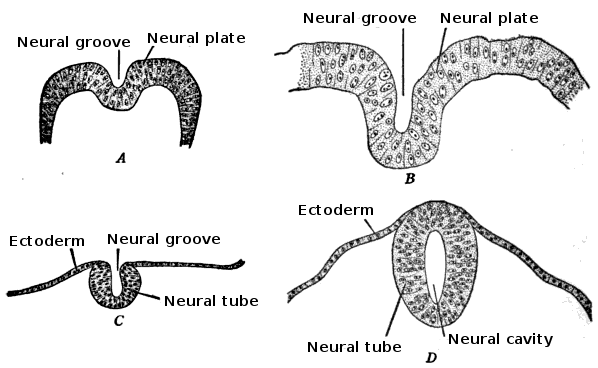
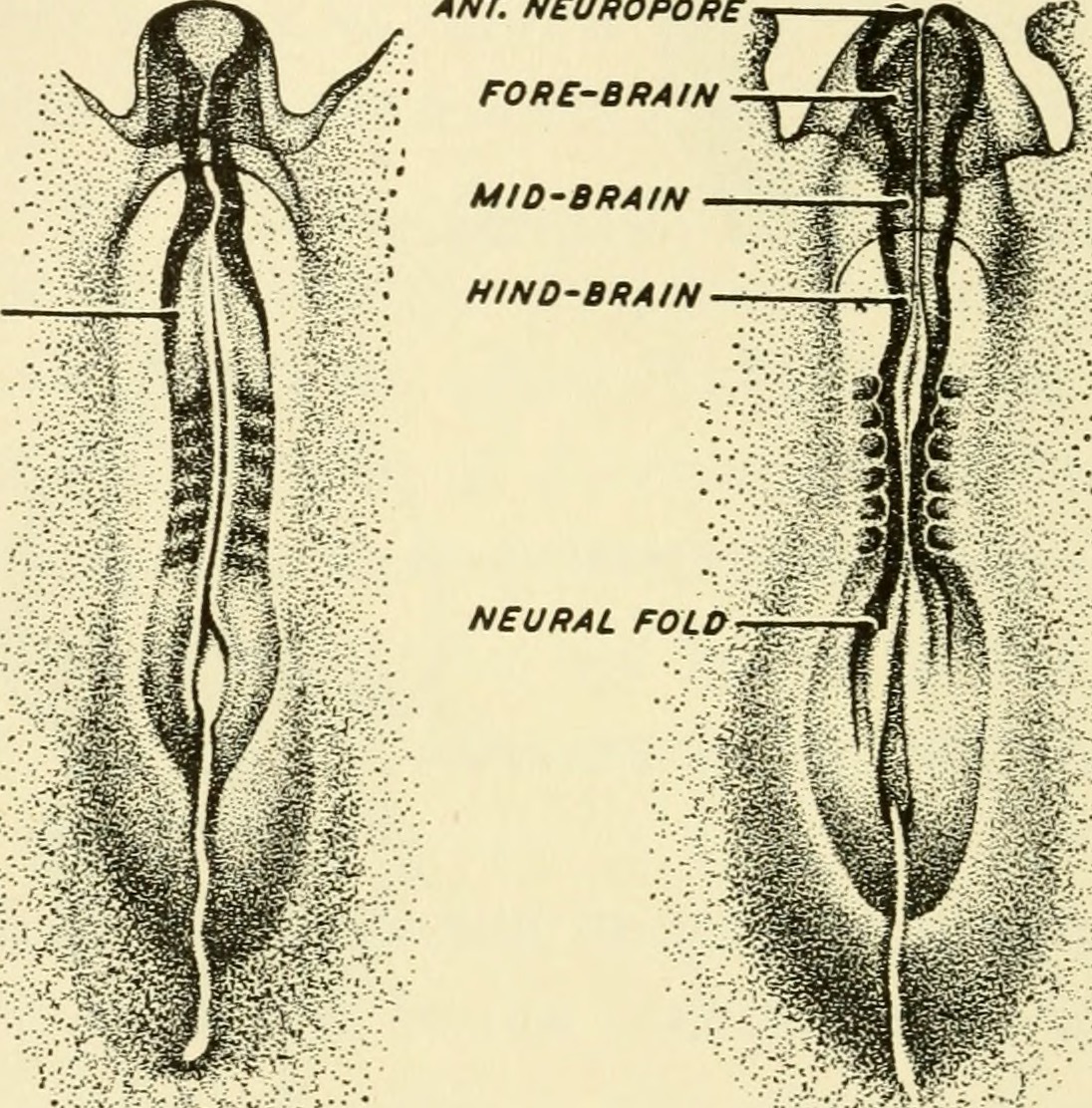
[1]
Moghadasi Boroujeni S, Koontz A, Tseropoulos G, Kerosuo L, Mehrotra P, Bajpai VK, Selvam SR, Lei P, Bronner ME, Andreadis ST. Neural crest stem cells from human epidermis of aged donors maintain their multipotency in vitro and in vivo. Scientific reports. 2019 Jul 5:9(1):9750. doi: 10.1038/s41598-019-46140-9. Epub 2019 Jul 5
[PubMed PMID: 31278326]
[2]
Duval N, Vaslin C, Barata TC, Frarma Y, Contremoulins V, Baudin X, Nedelec S, Ribes VC. BMP4 patterns Smad activity and generates stereotyped cell fate organization in spinal organoids. Development (Cambridge, England). 2019 Jul 25:146(14):. pii: dev175430. doi: 10.1242/dev.175430. Epub 2019 Jul 25
[PubMed PMID: 31239243]
[3]
Papaioannou E, Yánez DC, Ross S, Lau CI, Solanki A, Chawda MM, Virasami A, Ranz I, Ono M, O'Shaughnessy RFL, Crompton T. Sonic Hedgehog signaling limits atopic dermatitis via Gli2-driven immune regulation. The Journal of clinical investigation. 2019 Jul 2:129(8):3153-3170. doi: 10.1172/JCI125170. Epub 2019 Jul 2
[PubMed PMID: 31264977]
[4]
Severino M, Huisman TAGM. Posterior Fossa Malformations. Neuroimaging clinics of North America. 2019 Aug:29(3):367-383. doi: 10.1016/j.nic.2019.03.008. Epub 2019 May 2
[PubMed PMID: 31256860]
[5]
Durcanova B, Appleton J, Gurijala N, Belov V, Giffenig P, Moeller E, Hogan M, Lee F, Papisov M. The Configuration of the Perivascular System Transporting Macromolecules in the CNS. Frontiers in neuroscience. 2019:13():511. doi: 10.3389/fnins.2019.00511. Epub 2019 May 28
[PubMed PMID: 31191221]
[6]
Badaloni A, Casoni F, Croci L, Chiara F, Bizzoca A, Gennarini G, Cremona O, Hawkes R, Consalez GG. Dynamic Expression and New Functions of Early B Cell Factor 2 in Cerebellar Development. Cerebellum (London, England). 2019 Dec:18(6):999-1010. doi: 10.1007/s12311-019-01051-3. Epub
[PubMed PMID: 31273610]
[7]
Morimoto Y, Tamura R, Ohara K, Kosugi K, Oishi Y, Kuranari Y, Yoshida K, Toda M. Prognostic significance of VEGF receptors expression on the tumor cells in skull base chordoma. Journal of neuro-oncology. 2019 Aug:144(1):65-77. doi: 10.1007/s11060-019-03221-z. Epub 2019 Jun 25
[PubMed PMID: 31240525]
[8]
Spasojević G, Vujmilović S, Vujković Z, Gajanin R, Malobabić S, Ponorac N, Preradović L. Internal carotid and vertebral arteries diameters and their interrelationships to sex and left/right side. Folia morphologica. 2020:79(2):219-225. doi: 10.5603/FM.a2019.0071. Epub 2019 Jul 1
[PubMed PMID: 31257563]
[9]
Stojanović NN, Kostić A, Mitić R, Berilažić L, Radisavljević M. Association between Circle of Willis Configuration and Rupture of Cerebral Aneurysms. Medicina (Kaunas, Lithuania). 2019 Jul 3:55(7):. doi: 10.3390/medicina55070338. Epub 2019 Jul 3
[PubMed PMID: 31277348]
[10]
Anello MG, Miao TL, Pandey SK, Mandzia JL. Rare Bilateral Caudate Infarction in a Patient with a Common Circle of Willis Variant. The Canadian journal of neurological sciences. Le journal canadien des sciences neurologiques. 2019 Sep:46(5):593-594. doi: 10.1017/cjn.2019.233. Epub 2019 Jul 31
[PubMed PMID: 31230603]
[11]
Tamura R, Yoshida K, Toda M. Current understanding of lymphatic vessels in the central nervous system. Neurosurgical review. 2020 Aug:43(4):1055-1064. doi: 10.1007/s10143-019-01133-0. Epub 2019 Jun 18
[PubMed PMID: 31209659]
Level 3 (low-level) evidence
[12]
Son A, O'Donnell TF Jr, Izhakoff J, Gaebler JA, Niecko T, Iafrati MA. Lymphedema-associated comorbidities and treatment gap. Journal of vascular surgery. Venous and lymphatic disorders. 2019 Sep:7(5):724-730. doi: 10.1016/j.jvsv.2019.02.015. Epub 2019 Jun 24
[PubMed PMID: 31248833]
[13]
Yamaguchi Y, Miyazawa H, Miura M. Neural tube closure and embryonic metabolism. Congenital anomalies. 2017 Sep:57(5):134-137. doi: 10.1111/cga.12219. Epub 2017 May 31
[PubMed PMID: 28295633]
[14]
Sisman Y,Thomsen RH,Vestermark V,Krebs L, [Folate deficiency as a differential diagnosis to severe pre-eclampsia]. Ugeskrift for laeger. 2019 Jul 1;
[PubMed PMID: 31267942]
[15]
Goto T, Sakai T, Sato N, Katoh S, Sairyo K. An Adolescent Athlete with Low Back Pain Associated with Spina Bifida Occulta at the Thoracolumbar Junction : A Case Report. The journal of medical investigation : JMI. 2019:66(1.2):199-200. doi: 10.2152/jmi.66.199. Epub
[PubMed PMID: 31064941]
Level 3 (low-level) evidence
[17]
Struksnæs C,Blaas HK,Vogt C, Autopsy Findings of Central Nervous System Anomalies in Intact Fetuses Following Termination of Pregnancy After Prenatal Ultrasound Diagnosis. Pediatric and developmental pathology : the official journal of the Society for Pediatric Pathology and the Paediatric Pathology Society. 2019 Jun 29;
[PubMed PMID: 31256740]
[18]
Naikwadi A, Rege R, Hameed S. Antenatal sonographic diagnosis of semilobar holoprosencephaly with associated cleft lip and palate. BJR case reports. 2019 Feb:5(1):20180013. doi: 10.1259/bjrcr.20180013. Epub 2018 Oct 11
[PubMed PMID: 31131114]
Level 3 (low-level) evidence
[19]
Calloni SF,Caschera L,Triulzi FM, Disorders of Ventral Induction/Spectrum of Holoprosencephaly. Neuroimaging clinics of North America. 2019 Aug;
[PubMed PMID: 31256862]