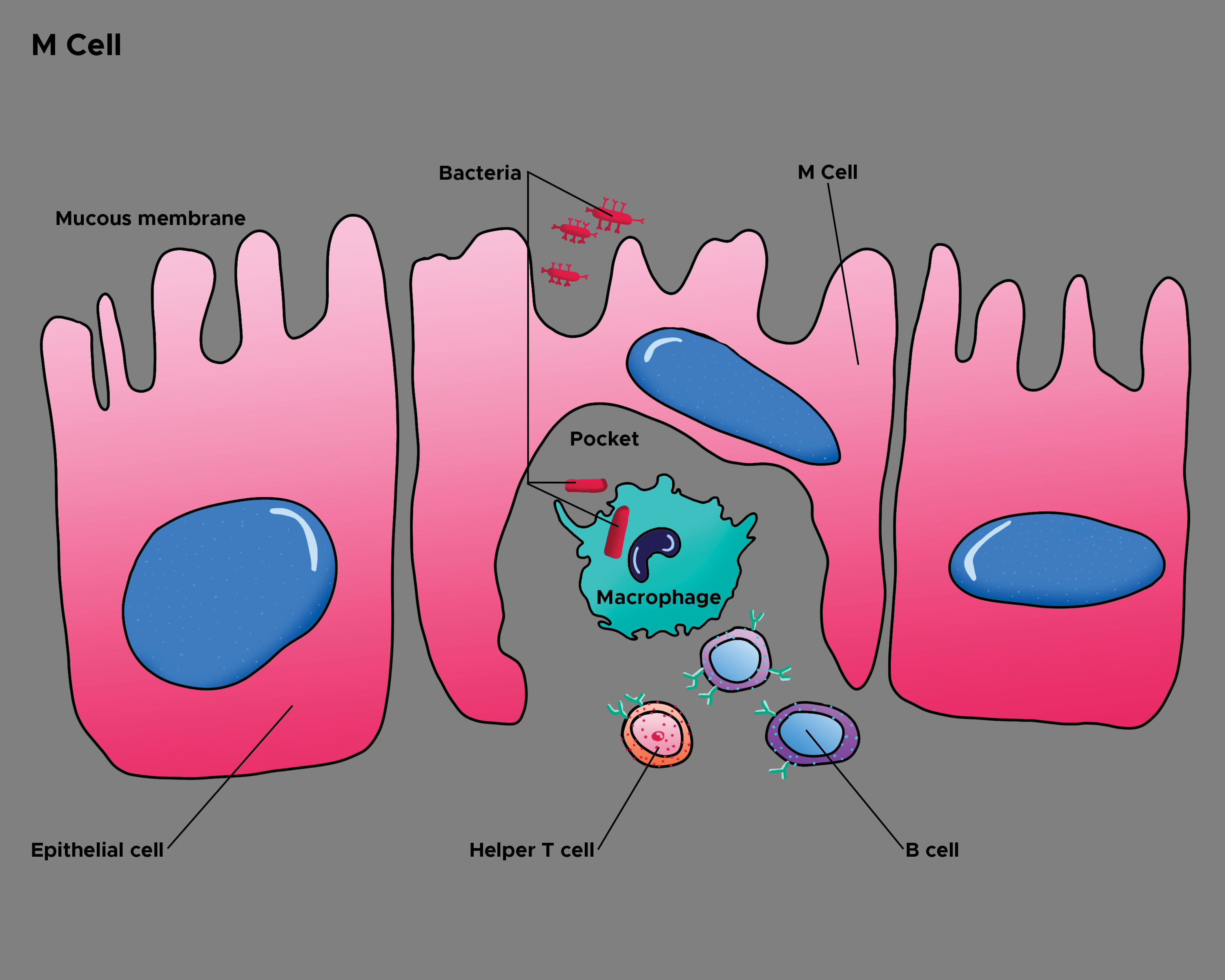
[1]
Corr SC, Gahan CC, Hill C. M-cells: origin, morphology and role in mucosal immunity and microbial pathogenesis. FEMS immunology and medical microbiology. 2008 Jan:52(1):2-12
[PubMed PMID: 18081850]
[2]
Kraehenbuhl JP, Corbett M. Immunology. Keeping the gut microflora at bay. Science (New York, N.Y.). 2004 Mar 12:303(5664):1624-5
[PubMed PMID: 15016988]
[3]
Gebert A, Rothkötter HJ, Pabst R. M cells in Peyer's patches of the intestine. International review of cytology. 1996:167():91-159
[PubMed PMID: 8768493]
[4]
Owen RL, Uptake and transport of intestinal macromolecules and microorganisms by M cells in Peyer's patches--a personal and historical perspective. Seminars in immunology. 1999 Jun
[PubMed PMID: 10381861]
Level 3 (low-level) evidence
[5]
Kucharzik T, Lügering N, Rautenberg K, Lügering A, Schmidt MA, Stoll R, Domschke W. Role of M cells in intestinal barrier function. Annals of the New York Academy of Sciences. 2000:915():171-83
[PubMed PMID: 11193574]
[6]
Rosner AJ, Keren DF. Demonstration of M cells in the specialized follicle-associated epithelium overlying isolated lymphoid follicles in the gut. Journal of leukocyte biology. 1984 Apr:35(4):397-404
[PubMed PMID: 6200555]
[7]
Mabbott NA, Kobayashi A, Sehgal A, Bradford BM, Pattison M, Donaldson DS. Aging and the mucosal immune system in the intestine. Biogerontology. 2015 Apr:16(2):133-45. doi: 10.1007/s10522-014-9498-z. Epub 2014 Apr 5
[PubMed PMID: 24705962]
[8]
Sehgal A,Kobayashi A,Donaldson DS,Mabbott NA, c-Rel is dispensable for the differentiation and functional maturation of M cells in the follicle-associated epithelium. Immunobiology. 2017 Feb
[PubMed PMID: 27663963]
[9]
Rouch JD, Scott A, Lei NY, Solorzano-Vargas RS, Wang J, Hanson EM, Kobayashi M, Lewis M, Stelzner MG, Dunn JC, Eckmann L, Martín MG. Development of Functional Microfold (M) Cells from Intestinal Stem Cells in Primary Human Enteroids. PloS one. 2016:11(1):e0148216. doi: 10.1371/journal.pone.0148216. Epub 2016 Jan 28
[PubMed PMID: 26820624]
[10]
Rios D, Wood MB, Li J, Chassaing B, Gewirtz AT, Williams IR. Antigen sampling by intestinal M cells is the principal pathway initiating mucosal IgA production to commensal enteric bacteria. Mucosal immunology. 2016 Jul:9(4):907-16. doi: 10.1038/mi.2015.121. Epub 2015 Nov 25
[PubMed PMID: 26601902]
[11]
Schmucker DL, Thoreux K, Owen RL. Aging impairs intestinal immunity. Mechanisms of ageing and development. 2001 Sep 15:122(13):1397-411
[PubMed PMID: 11470129]
[12]
Gullberg E,Söderholm JD, Peyer's patches and M cells as potential sites of the inflammatory onset in Crohn's disease. Annals of the New York Academy of Sciences. 2006 Aug
[PubMed PMID: 17057202]
[13]
Owen RL. M cells as portals of entry for HIV. Pathobiology : journal of immunopathology, molecular and cellular biology. 1998:66(3-4):141-4
[PubMed PMID: 9693315]
[14]
Wang M, Gao Z, Zhang Z, Pan L, Zhang Y. Roles of M cells in infection and mucosal vaccines. Human vaccines & immunotherapeutics. 2014:10(12):3544-51. doi: 10.4161/hv.36174. Epub
[PubMed PMID: 25483705]
[15]
Takahashi K, Yano A, Watanabe S, Langella P, Bermúdez-Humarán LG, Inoue N. M cell-targeting strategy enhances systemic and mucosal immune responses induced by oral administration of nuclease-producing L. lactis. Applied microbiology and biotechnology. 2018 Dec:102(24):10703-10711. doi: 10.1007/s00253-018-9427-1. Epub 2018 Oct 11
[PubMed PMID: 30310964]
[16]
Azizi A,Kumar A,Diaz-Mitoma F,Mestecky J, Enhancing oral vaccine potency by targeting intestinal M cells. PLoS pathogens. 2010 Nov 11
[PubMed PMID: 21085599]