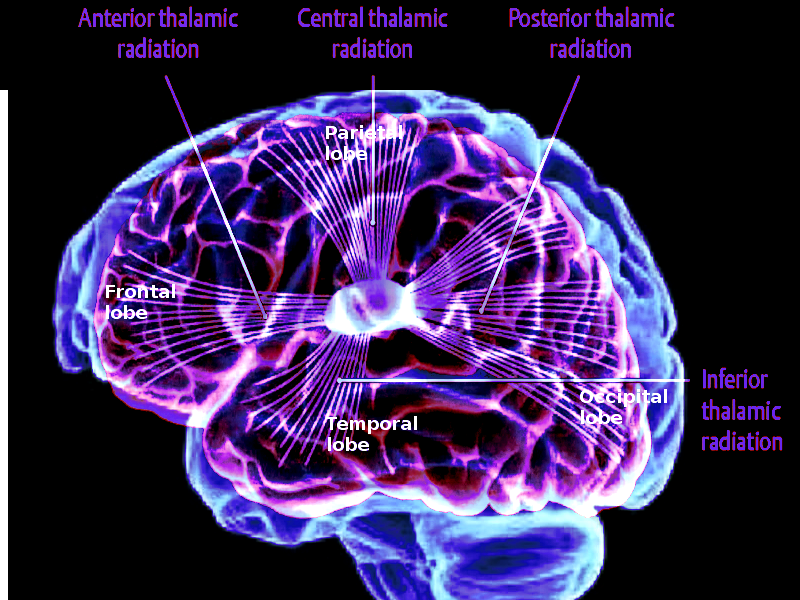
[1]
Jones EG. Thalamic circuitry and thalamocortical synchrony. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 2002 Dec 29:357(1428):1659-73
[PubMed PMID: 12626002]
[2]
Zhang Y, Zhang J, Oishi K, Faria AV, Jiang H, Li X, Akhter K, Rosa-Neto P, Pike GB, Evans A, Toga AW, Woods R, Mazziotta JC, Miller MI, van Zijl PC, Mori S. Atlas-guided tract reconstruction for automated and comprehensive examination of the white matter anatomy. NeuroImage. 2010 Oct 1:52(4):1289-301. doi: 10.1016/j.neuroimage.2010.05.049. Epub 2010 May 24
[PubMed PMID: 20570617]
[3]
Woodward ND, Karbasforoushan H, Heckers S. Thalamocortical dysconnectivity in schizophrenia. The American journal of psychiatry. 2012 Oct:169(10):1092-9. doi: 10.1176/appi.ajp.2012.12010056. Epub
[PubMed PMID: 23032387]
[4]
Scheibel ME, Scheibel AB. Structural organization of nonspecific thalamic nuclei and their projection toward cortex. Brain research. 1967 Sep:6(1):60-94
[PubMed PMID: 4964024]
[5]
Briggs F, Usrey WM. Emerging views of corticothalamic function. Current opinion in neurobiology. 2008 Aug:18(4):403-7. doi: 10.1016/j.conb.2008.09.002. Epub 2008 Oct 6
[PubMed PMID: 18805486]
Level 3 (low-level) evidence
[7]
Palmer SL, Noctor SC, Jablonska B, Juliano SL. Laminar specific alterations of thalamocortical projections in organotypic cultures following layer 4 disruption in ferret somatosensory cortex. The European journal of neuroscience. 2001 Apr:13(8):1559-71
[PubMed PMID: 11328350]
[8]
Kharazia VN, Weinberg RJ. Glutamate in thalamic fibers terminating in layer IV of primary sensory cortex. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1994 Oct:14(10):6021-32
[PubMed PMID: 7931559]
[9]
Krsnik Ž, Majić V, Vasung L, Huang H, Kostović I. Growth of Thalamocortical Fibers to the Somatosensory Cortex in the Human Fetal Brain. Frontiers in neuroscience. 2017:11():233. doi: 10.3389/fnins.2017.00233. Epub 2017 Apr 27
[PubMed PMID: 28496398]
[10]
Molnár Z. Development and evolution of thalamocortical interactions. European journal of morphology. 2000 Dec:38(5):313-20
[PubMed PMID: 11151044]
[12]
Freeman W, Watts JW. Prefrontal Lobotomy: The Surgical Relief of Mental Pain. Bulletin of the New York Academy of Medicine. 1942 Dec:18(12):794-812
[PubMed PMID: 19312295]
[13]
Zhou J, Liu X, Song W, Yang Y, Zhao Z, Ling F, Hudetz AG, Li SJ. Specific and nonspecific thalamocortical functional connectivity in normal and vegetative states. Consciousness and cognition. 2011 Jun:20(2):257-68. doi: 10.1016/j.concog.2010.08.003. Epub 2010 Nov 13
[PubMed PMID: 21078562]
[14]
Laureys S, Faymonville ME, Luxen A, Lamy M, Franck G, Maquet P. Restoration of thalamocortical connectivity after recovery from persistent vegetative state. Lancet (London, England). 2000 May 20:355(9217):1790-1
[PubMed PMID: 10832834]
[15]
Kostopoulos GK. Involvement of the thalamocortical system in epileptic loss of consciousness. Epilepsia. 2001:42 Suppl 3():13-9
[PubMed PMID: 11520316]
[16]
DeLong MR, Wichmann T. Basal Ganglia Circuits as Targets for Neuromodulation in Parkinson Disease. JAMA neurology. 2015 Nov:72(11):1354-60. doi: 10.1001/jamaneurol.2015.2397. Epub
[PubMed PMID: 26409114]
[17]
Pulsipher DT, Seidenberg M, Guidotti L, Tuchscherer VN, Morton J, Sheth RD, Hermann B. Thalamofrontal circuitry and executive dysfunction in recent-onset juvenile myoclonic epilepsy. Epilepsia. 2009 May:50(5):1210-9. doi: 10.1111/j.1528-1167.2008.01952.x. Epub 2009 Jan 17
[PubMed PMID: 19183226]
[18]
Wang R, Fan Q, Zhang Z, Chen Y, Zhu Y, Li Y. Anterior thalamic radiation structural and metabolic changes in obsessive-compulsive disorder: A combined DTI-MRS study. Psychiatry research. Neuroimaging. 2018 Jul 30:277():39-44. doi: 10.1016/j.pscychresns.2018.05.004. Epub 2018 May 12
[PubMed PMID: 29807209]
[19]
Frieling H, Fischer J, Wilhelm J, Engelhorn T, Bleich S, Hillemacher T, Dörfler A, Kornhuber J, de Zwaan M, Peschel T. Microstructural abnormalities of the posterior thalamic radiation and the mediodorsal thalamic nuclei in females with anorexia nervosa--a voxel based diffusion tensor imaging (DTI) study. Journal of psychiatric research. 2012 Sep:46(9):1237-42. doi: 10.1016/j.jpsychires.2012.06.005. Epub 2012 Jul 4
[PubMed PMID: 22770509]
[20]
Mamah D, Conturo TE, Harms MP, Akbudak E, Wang L, McMichael AR, Gado MH, Barch DM, Csernansky JG. Anterior thalamic radiation integrity in schizophrenia: a diffusion-tensor imaging study. Psychiatry research. 2010 Aug 30:183(2):144-50. doi: 10.1016/j.pscychresns.2010.04.013. Epub 2010 Jul 8
[PubMed PMID: 20619618]
[21]
Nair A, Treiber JM, Shukla DK, Shih P, Müller RA. Impaired thalamocortical connectivity in autism spectrum disorder: a study of functional and anatomical connectivity. Brain : a journal of neurology. 2013 Jun:136(Pt 6):1942-55. doi: 10.1093/brain/awt079. Epub
[PubMed PMID: 23739917]
[22]
Magon S, May A, Stankewitz A, Goadsby PJ, Tso AR, Ashina M, Amin FM, Seifert CL, Chakravarty MM, Müller J, Sprenger T. Morphological Abnormalities of Thalamic Subnuclei in Migraine: A Multicenter MRI Study at 3 Tesla. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2015 Oct 7:35(40):13800-6. doi: 10.1523/JNEUROSCI.2154-15.2015. Epub
[PubMed PMID: 26446230]
[23]
Taylor JP, Firbank M, Barnett N, Pearce S, Livingstone A, Mosimann U, Eyre J, McKeith IG, O'Brien JT. Visual hallucinations in dementia with Lewy bodies: transcranial magnetic stimulation study. The British journal of psychiatry : the journal of mental science. 2011 Dec:199(6):492-500. doi: 10.1192/bjp.bp.110.090373. Epub 2011 Oct 20
[PubMed PMID: 22016436]