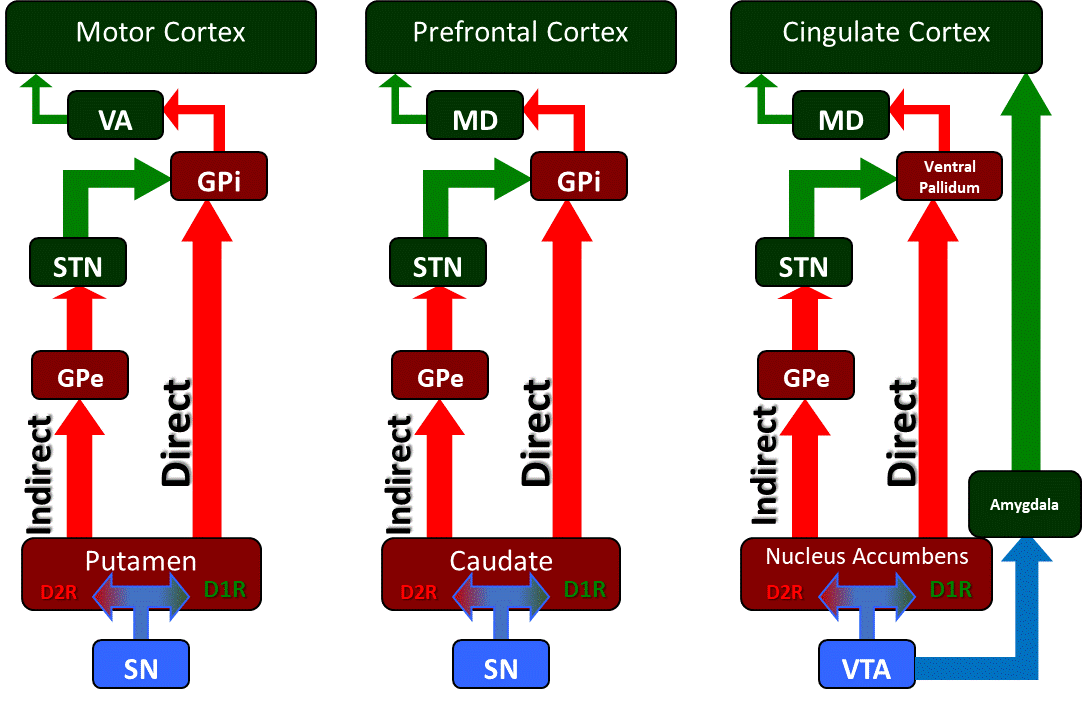
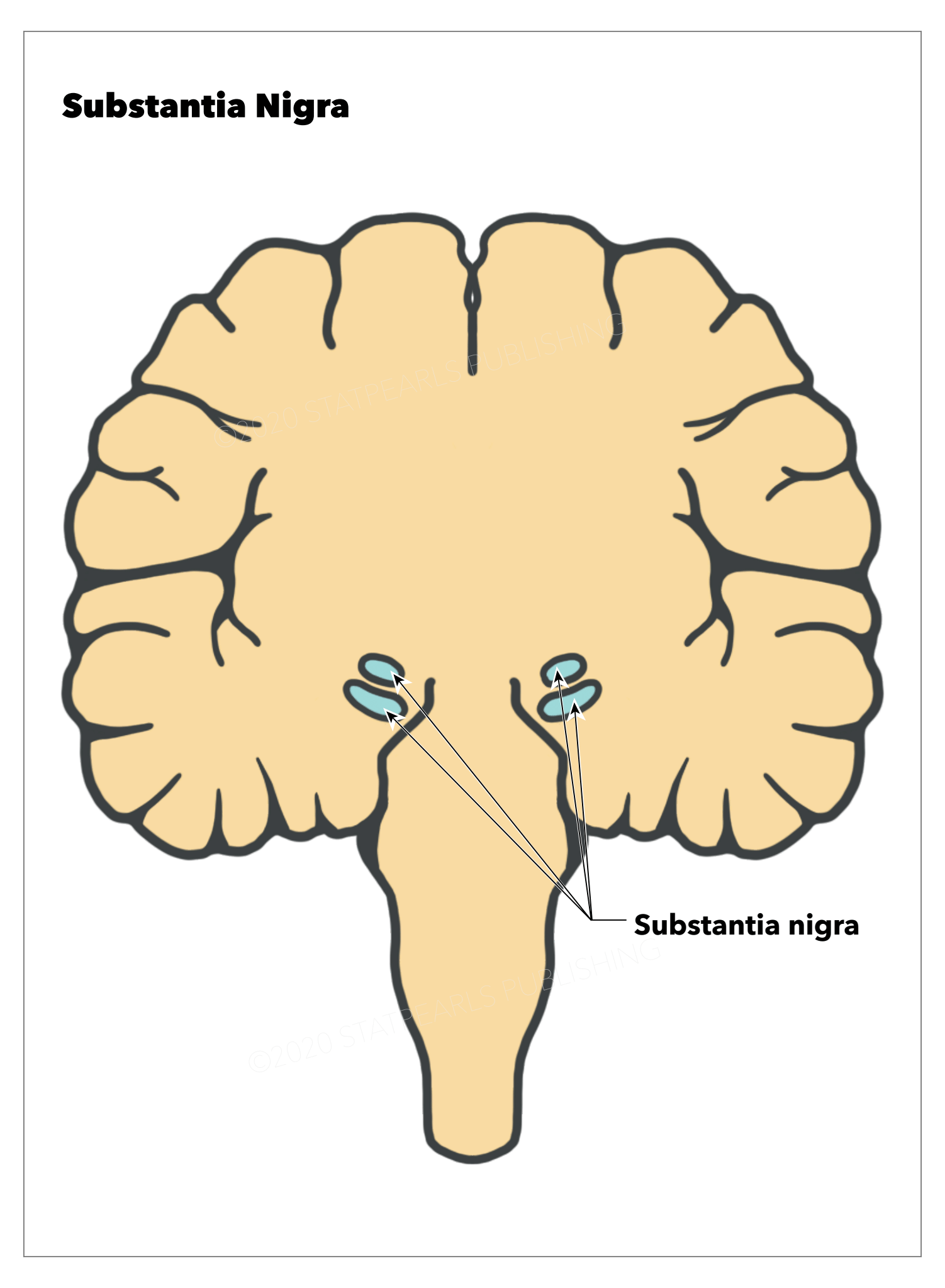
[1]
Hodge GK, Butcher LL. Pars compacta of the substantia nigra modulates motor activity but is not involved importantly in regulating food and water intake. Naunyn-Schmiedeberg's archives of pharmacology. 1980 Aug:313(1):51-67
[PubMed PMID: 7207636]
[2]
Fabbri M, Reimão S, Carvalho M, Nunes RG, Abreu D, Guedes LC, Bouça R, Lobo PP, Godinho C, Coelho M, Gonçalves NC, Rosa MM, Antonini A, Ferreira JJ. Substantia Nigra Neuromelanin as an Imaging Biomarker of Disease Progression in Parkinson's Disease. Journal of Parkinson's disease. 2017:7(3):491-501. doi: 10.3233/JPD-171135. Epub
[PubMed PMID: 28671143]
[3]
Galtieri DJ, Estep CM, Wokosin DL, Traynelis S, Surmeier DJ. Pedunculopontine glutamatergic neurons control spike patterning in substantia nigra dopaminergic neurons. eLife. 2017 Oct 5:6():. doi: 10.7554/eLife.30352. Epub 2017 Oct 5
[PubMed PMID: 28980939]
[4]
Freeman TB,Spence MS,Boss BD,Spector DH,Strecker RE,Olanow CW,Kordower JH, Development of dopaminergic neurons in the human substantia nigra. Experimental neurology. 1991 Sep
[PubMed PMID: 1680741]
[5]
Aubert I, Brana C, Pellevoisin C, Giros B, Caille I, Carles D, Vital C, Bloch B. Molecular anatomy of the development of the human substantia nigra. The Journal of comparative neurology. 1997 Mar 3:379(1):72-87
[PubMed PMID: 9057113]
Level 2 (mid-level) evidence
[6]
Altman J, Bayer SA. Development of the brain stem in the rat. V. Thymidine-radiographic study of the time of origin of neurons in the midbrain tegmentum. The Journal of comparative neurology. 1981 Jun 1:198(4):677-716
[PubMed PMID: 7251936]
Level 2 (mid-level) evidence
[7]
Nunes I, Tovmasian LT, Silva RM, Burke RE, Goff SP. Pitx3 is required for development of substantia nigra dopaminergic neurons. Proceedings of the National Academy of Sciences of the United States of America. 2003 Apr 1:100(7):4245-50
[PubMed PMID: 12655058]
[8]
Le W,Zhang L,Xie W,Li S,Dani JA, Pitx3 deficiency produces decreased dopamine signaling and induces motor deficits in Pitx3(-/-) mice. Neurobiology of aging. 2015 Dec
[PubMed PMID: 26363812]
[9]
Villaescusa JC, Li B, Toledo EM, Rivetti di Val Cervo P, Yang S, Stott SR, Kaiser K, Islam S, Gyllborg D, Laguna-Goya R, Landreh M, Lönnerberg P, Falk A, Bergman T, Barker RA, Linnarsson S, Selleri L, Arenas E. A PBX1 transcriptional network controls dopaminergic neuron development and is impaired in Parkinson's disease. The EMBO journal. 2016 Sep 15:35(18):1963-78. doi: 10.15252/embj.201593725. Epub 2016 Jun 28
[PubMed PMID: 27354364]
[10]
Valente EM, Abou-Sleiman PM, Caputo V, Muqit MM, Harvey K, Gispert S, Ali Z, Del Turco D, Bentivoglio AR, Healy DG, Albanese A, Nussbaum R, González-Maldonado R, Deller T, Salvi S, Cortelli P, Gilks WP, Latchman DS, Harvey RJ, Dallapiccola B, Auburger G, Wood NW. Hereditary early-onset Parkinson's disease caused by mutations in PINK1. Science (New York, N.Y.). 2004 May 21:304(5674):1158-60
[PubMed PMID: 15087508]
Level 2 (mid-level) evidence
[11]
Kitada T, Asakawa S, Hattori N, Matsumine H, Yamamura Y, Minoshima S, Yokochi M, Mizuno Y, Shimizu N. Mutations in the parkin gene cause autosomal recessive juvenile parkinsonism. Nature. 1998 Apr 9:392(6676):605-8
[PubMed PMID: 9560156]
[12]
Abou-Sleiman PM, Hanna MG, Wood NW. Genetic association studies of complex neurological diseases. Journal of neurology, neurosurgery, and psychiatry. 2006 Dec:77(12):1302-4
[PubMed PMID: 17110744]
[13]
Mata IF, Wedemeyer WJ, Farrer MJ, Taylor JP, Gallo KA. LRRK2 in Parkinson's disease: protein domains and functional insights. Trends in neurosciences. 2006 May:29(5):286-93
[PubMed PMID: 16616379]
[14]
Manosalva HA, Pio F, Jeerakathil T, Saqqur M, Camicioli R, Suchowersky O. Vascular Parkinsonism in a Tertiary Care Stroke Prevention Clinic and the Development of a New Screening Strategy. Journal of stroke and cerebrovascular diseases : the official journal of National Stroke Association. 2018 Jan:27(1):153-161. doi: 10.1016/j.jstrokecerebrovasdis.2017.08.020. Epub 2017 Oct 3
[PubMed PMID: 28986199]
[15]
Brydon L, Harrison NA, Walker C, Steptoe A, Critchley HD. Peripheral inflammation is associated with altered substantia nigra activity and psychomotor slowing in humans. Biological psychiatry. 2008 Jun 1:63(11):1022-9. doi: 10.1016/j.biopsych.2007.12.007. Epub 2008 Feb 1
[PubMed PMID: 18242584]
[16]
Yang L, Wang H, Liu L, Xie A. The Role of Insulin/IGF-1/PI3K/Akt/GSK3β Signaling in Parkinson's Disease Dementia. Frontiers in neuroscience. 2018:12():73. doi: 10.3389/fnins.2018.00073. Epub 2018 Feb 20
[PubMed PMID: 29515352]
[17]
Langston JW, Irwin I, Langston EB, Forno LS. 1-Methyl-4-phenylpyridinium ion (MPP+): identification of a metabolite of MPTP, a toxin selective to the substantia nigra. Neuroscience letters. 1984 Jul 13:48(1):87-92
[PubMed PMID: 6332288]
Level 3 (low-level) evidence
[18]
Gash DM,Rutland K,Hudson NL,Sullivan PG,Bing G,Cass WA,Pandya JD,Liu M,Choi DY,Hunter RL,Gerhardt GA,Smith CD,Slevin JT,Prince TS, Trichloroethylene: Parkinsonism and complex 1 mitochondrial neurotoxicity. Annals of neurology. 2008 Feb
[PubMed PMID: 18157908]
[19]
Tanner CM, Kamel F, Ross GW, Hoppin JA, Goldman SM, Korell M, Marras C, Bhudhikanok GS, Kasten M, Chade AR, Comyns K, Richards MB, Meng C, Priestley B, Fernandez HH, Cambi F, Umbach DM, Blair A, Sandler DP, Langston JW. Rotenone, paraquat, and Parkinson's disease. Environmental health perspectives. 2011 Jun:119(6):866-72. doi: 10.1289/ehp.1002839. Epub 2011 Jan 26
[PubMed PMID: 21269927]
Level 3 (low-level) evidence
[20]
Stenslik MJ, Potts LF, Sonne JW, Cass WA, Turchan-Cholewo J, Pomerleau F, Huettl P, Ai Y, Gash DM, Gerhardt GA, Bradley LH. Methodology and effects of repeated intranasal delivery of DNSP-11 in a rat model of Parkinson's disease. Journal of neuroscience methods. 2015 Aug 15:251():120-9. doi: 10.1016/j.jneumeth.2015.05.006. Epub 2015 May 18
[PubMed PMID: 25999268]