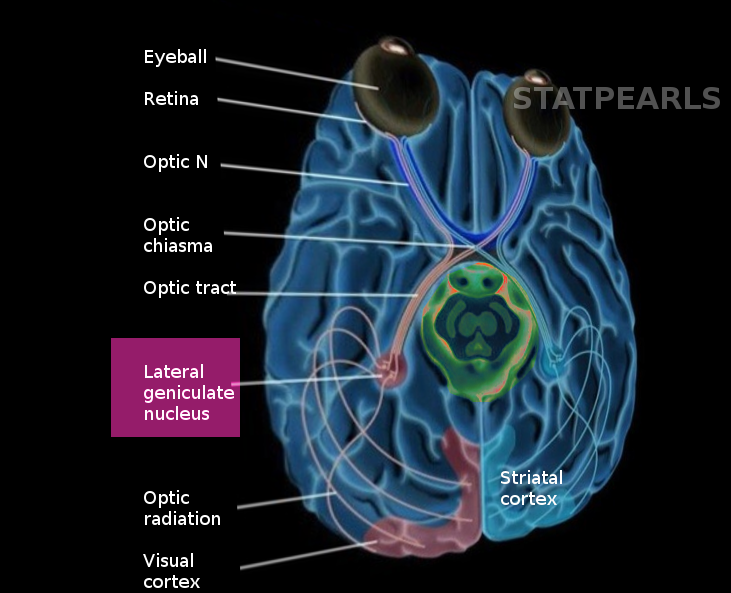
[1]
Schmid MC, Mrowka SW, Turchi J, Saunders RC, Wilke M, Peters AJ, Ye FQ, Leopold DA. Blindsight depends on the lateral geniculate nucleus. Nature. 2010 Jul 15:466(7304):373-7. doi: 10.1038/nature09179. Epub 2010 Jun 23
[PubMed PMID: 20574422]
[2]
Sherman SM, Guillery RW. The role of the thalamus in the flow of information to the cortex. Philosophical transactions of the Royal Society of London. Series B, Biological sciences. 2002 Dec 29:357(1428):1695-708
[PubMed PMID: 12626004]
[3]
Yoshida M, Sasa M, Takaori S. Serotonin-mediated inhibition from dorsal raphe nucleus of neurons in dorsal lateral geniculate and thalamic reticular nuclei. Brain research. 1984 Jan 2:290(1):95-105
[PubMed PMID: 6692141]
[4]
Krueger J, Disney AA. Structure and function of dual-source cholinergic modulation in early vision. The Journal of comparative neurology. 2019 Feb 15:527(3):738-750. doi: 10.1002/cne.24590. Epub 2018 Dec 20
[PubMed PMID: 30520037]
Level 2 (mid-level) evidence
[5]
Holdefer RN, Jacobs BL. Phasic stimulation of the locus coeruleus: effects on activity in the lateral geniculate nucleus. Experimental brain research. 1994:100(3):444-52
[PubMed PMID: 7813682]
[6]
Brückner G, Morawski M, Arendt T. Aggrecan-based extracellular matrix is an integral part of the human basal ganglia circuit. Neuroscience. 2008 Jan 24:151(2):489-504
[PubMed PMID: 18055126]
[7]
Lendvai D, Morawski M, Brückner G, Négyessy L, Baksa G, Glasz T, Patonay L, Matthews RT, Arendt T, Alpár A. Perisynaptic aggrecan-based extracellular matrix coats in the human lateral geniculate body devoid of perineuronal nets. Journal of neuroscience research. 2012 Feb:90(2):376-87. doi: 10.1002/jnr.22761. Epub 2011 Sep 30
[PubMed PMID: 21959900]
[8]
Moore RY, Card JP. Intergeniculate leaflet: an anatomically and functionally distinct subdivision of the lateral geniculate complex. The Journal of comparative neurology. 1994 Jun 15:344(3):403-30
[PubMed PMID: 8063960]
Level 2 (mid-level) evidence
[9]
Muscat L, Morin LP. Intergeniculate leaflet: contributions to photic and non-photic responsiveness of the hamster circadian system. Neuroscience. 2006 Jun 19:140(1):305-20
[PubMed PMID: 16549274]
[10]
Cipolla-Neto J, Bartol I, Seraphim PM, Afeche SC, Scialfa JH, Peraçoli AM. The effects of lesions of the thalamic intergeniculate leaflet on the pineal metabolism. Brain research. 1995 Sep 11:691(1-2):133-41
[PubMed PMID: 8590045]
[11]
Glees P, le Gros Clark WE. The termination of optic fibres in the lateral geniculate body of the monkey. Journal of anatomy. 1941 Apr:75(Pt 3):295-308.3
[PubMed PMID: 17104862]
[12]
O'Connor DH, Fukui MM, Pinsk MA, Kastner S. Attention modulates responses in the human lateral geniculate nucleus. Nature neuroscience. 2002 Nov:5(11):1203-9
[PubMed PMID: 12379861]
[13]
Rathbun DL, Warland DK, Usrey WM. Spike timing and information transmission at retinogeniculate synapses. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2010 Oct 13:30(41):13558-66. doi: 10.1523/JNEUROSCI.0909-10.2010. Epub
[PubMed PMID: 20943897]
[14]
Dan Y, Atick JJ, Reid RC. Efficient coding of natural scenes in the lateral geniculate nucleus: experimental test of a computational theory. The Journal of neuroscience : the official journal of the Society for Neuroscience. 1996 May 15:16(10):3351-62
[PubMed PMID: 8627371]
[15]
Dougherty K, Schmid MC, Maier A. Binocular response modulation in the lateral geniculate nucleus. The Journal of comparative neurology. 2019 Feb 15:527(3):522-534. doi: 10.1002/cne.24417. Epub 2018 Mar 9
[PubMed PMID: 29473163]
Level 2 (mid-level) evidence
[16]
Cheong SK, Tailby C, Solomon SG, Martin PR. Cortical-like receptive fields in the lateral geniculate nucleus of marmoset monkeys. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2013 Apr 17:33(16):6864-76. doi: 10.1523/JNEUROSCI.5208-12.2013. Epub
[PubMed PMID: 23595745]
[17]
COOPER ER. The development of the human lateral geniculate body. Brain : a journal of neurology. 1945:68(3):222-39
[PubMed PMID: 21017988]
[18]
Khan AA, Wadhwa S, Bijlani V. Development of human lateral geniculate nucleus: an electron microscopic study. International journal of developmental neuroscience : the official journal of the International Society for Developmental Neuroscience. 1994 Nov:12(7):661-72
[PubMed PMID: 7900548]
[19]
Hevner RF. Development of connections in the human visual system during fetal mid-gestation: a DiI-tracing study. Journal of neuropathology and experimental neurology. 2000 May:59(5):385-92
[PubMed PMID: 10888368]
[20]
Hitchcock PF, Hickey TL. Prenatal development of the human lateral geniculate nucleus. The Journal of comparative neurology. 1980 Nov 15:194(2):395-411
[PubMed PMID: 7440807]
Level 2 (mid-level) evidence
[21]
Huberman AD, Stellwagen D, Chapman B. Decoupling eye-specific segregation from lamination in the lateral geniculate nucleus. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2002 Nov 1:22(21):9419-29
[PubMed PMID: 12417667]
[22]
Luco C, Hoppe A, Schweitzer M, Vicuña X, Fantin A. Visual field defects in vascular lesions of the lateral geniculate body. Journal of neurology, neurosurgery, and psychiatry. 1992 Jan:55(1):12-5
[PubMed PMID: 1548490]
[23]
Weyand TG. The multifunctional lateral geniculate nucleus. Reviews in the neurosciences. 2016 Feb:27(2):135-57. doi: 10.1515/revneuro-2015-0018. Epub
[PubMed PMID: 26479339]
[24]
Barnes GR, Li X, Thompson B, Singh KD, Dumoulin SO, Hess RF. Decreased gray matter concentration in the lateral geniculate nuclei in human amblyopes. Investigative ophthalmology & visual science. 2010 Mar:51(3):1432-8. doi: 10.1167/iovs.09-3931. Epub 2009 Oct 29
[PubMed PMID: 19875650]
[25]
Gupta N, Greenberg G, de Tilly LN, Gray B, Polemidiotis M, Yücel YH. Atrophy of the lateral geniculate nucleus in human glaucoma detected by magnetic resonance imaging. The British journal of ophthalmology. 2009 Jan:93(1):56-60. doi: 10.1136/bjo.2008.138172. Epub 2008 Aug 12
[PubMed PMID: 18697810]
[26]
Dai H, Mu KT, Qi JP, Wang CY, Zhu WZ, Xia LM, Chen ZQ, Zhang H, Ai F, Morelli JN. Assessment of lateral geniculate nucleus atrophy with 3T MR imaging and correlation with clinical stage of glaucoma. AJNR. American journal of neuroradiology. 2011 Aug:32(7):1347-53. doi: 10.3174/ajnr.A2486. Epub 2011 Jul 14
[PubMed PMID: 21757515]
Level 2 (mid-level) evidence
[27]
Al-Abdulla NA, Portera-Cailliau C, Martin LJ. Occipital cortex ablation in adult rat causes retrograde neuronal death in the lateral geniculate nucleus that resembles apoptosis. Neuroscience. 1998 Sep:86(1):191-209
[PubMed PMID: 9692754]
[28]
Sizdahkhani S, Magill ST, McDermott MW. Intraventricular Meningioma Resection with Postoperative Ischemia of the Lateral Geniculate Nucleus. World neurosurgery. 2017 Oct:106():878-883. doi: 10.1016/j.wneu.2017.07.067. Epub 2017 Jul 19
[PubMed PMID: 28735134]
[29]
Gunderson CH, Hoyt WF. Geniculate hemianopia: incongruous homonymous field defects in two patients with partial lesions of the lateral geniculate nucleus. Journal of neurology, neurosurgery, and psychiatry. 1971 Feb:34(1):1-6
[PubMed PMID: 5551688]
[30]
Johnson RF, Smale L, Moore RY, Morin LP. Lateral geniculate lesions block circadian phase-shift responses to a benzodiazepine. Proceedings of the National Academy of Sciences of the United States of America. 1988 Jul:85(14):5301-4
[PubMed PMID: 3293053]
[31]
Rüb U, Del Turco D, Bürk K, Diaz GO, Auburger G, Mittelbronn M, Gierga K, Ghebremedhin E, Schultz C, Schöls L, Bohl J, Braak H, Deller T. Extended pathoanatomical studies point to a consistent affection of the thalamus in spinocerebellar ataxia type 2. Neuropathology and applied neurobiology. 2005 Apr:31(2):127-40
[PubMed PMID: 15771706]
[32]
Rüb U, Del Turco D, Del Tredici K, de Vos RA, Brunt ER, Reifenberger G, Seifried C, Schultz C, Auburger G, Braak H. Thalamic involvement in a spinocerebellar ataxia type 2 (SCA2) and a spinocerebellar ataxia type 3 (SCA3) patient, and its clinical relevance. Brain : a journal of neurology. 2003 Oct:126(Pt 10):2257-72
[PubMed PMID: 12847080]