Introduction
The brainstem is the structure that connects the cerebrum of the brain to the spinal cord and cerebellum. It is composed of three sections in descending order: the midbrain, pons, and medulla oblongata. It is responsible for many vital functions of life, such as breathing, consciousness, blood pressure, heart rate, and sleep. The brainstem contains many critical collections of white and grey matter. The grey matter within the brainstem consists of nerve cell bodies and forms many important brainstem nuclei. The white matter tracts of the brainstem include axons of nerves traversing their course to different structures; the axons originate from cell bodies located elsewhere within the central nervous system (CNS). Some of the white matter tract cell bodies are located within the brainstem as well. These tracts travel both to the brain (afferent) and from the brain (efferent), such as the somatosensory pathways and the corticospinal tracts, respectively. Ten of the twelve cranial nerves arise from their cranial nerve nuclei in the brainstem. Clinicians can localize lesions of the brainstem with a thorough knowledge of brainstem anatomy and functions. The following activity addresses the nuclei, tracts, and functions of the brainstem.
Structure and Function
The brainstem contains many different nuclei and tracts. This section overviews the major structures located in each area of the brainstem, followed by a summary of the course of the major pathways through the brainstem.
Midbrain
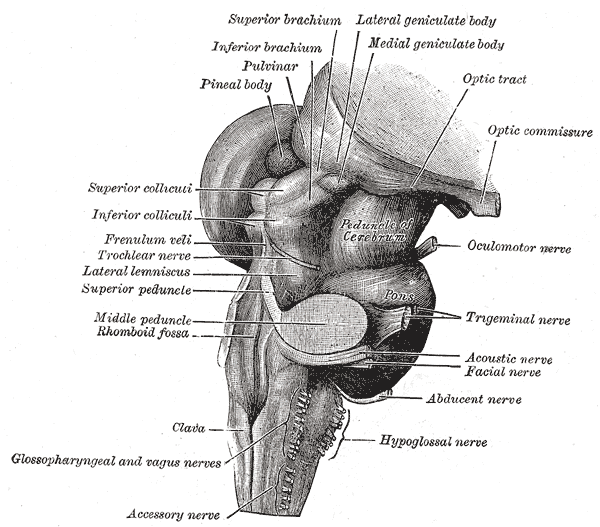
The midbrain serves as the connection between the pons and the diencephalon. It also connects posteriorly to the cerebellum via the superior cerebellar peduncles. The anterior part of the midbrain contains the crus cerebri with the interpeduncular fossa located between them. The crus cerebri carry motor cortical spinal fibers, corticonuclear fibers, and pontine fiber tracts. The midbrain contains the cerebral aqueduct centrally, which connects the third ventricle superiorly with the fourth ventricle inferiorly. The periaqueductal grey surrounds the cerebral aqueduct. The midbrain is separated relative to the cerebral aqueduct, with the posterior portion being the tectum (floor) and anterior to the aqueduct serving as the tegmentum (roof). The posterior surface of the midbrain contains the corpora quadrigemina, which are composed of bilateral superior colliculi and bilateral inferior colliculi. The superior colliculi are involved in visual reflexes such as saccadic eye movements. Each superior colliculus sends fibers to the corresponding lateral geniculate body and optic tract through the superior brachium. The inferior colliculi are involved in auditory processing and connect to their corresponding medial geniculate nuclei through the inferior brachium. Just inferior to the inferior colliculi at the posterior midline of the brainstem cranial nerve IV, the trochlear nerve emerges. The trochlear nerve is unique among cranial nerves as it is the only one to emerge from the posterior surface of the brainstem. The other cranial nerve that arises from the midbrain is cranial nerve III, the oculomotor nerve. The oculomotor nerve arises from the midbrain in the oculomotor sulcus on the medial surface of the crus cerebri, within the interpeduncular cistern.
The midbrain also includes many other important nuclei, including but not limited to the substantia nigra at the base of the midbrain, the red nucleus anterior medially at the level of the superior colliculus, and the dorsal raphe nucleus. The largest of the raphe nuclei are in the anterior midline of the periaqueductal grey, and the location of the ventral tegmental area is near the midline medial to the red nucleus. The substantia nigra contains dopaminergic neurons that help to regulate movement associated with the basal ganglia. The ventral tegmental area also contains dopaminergic neurons and plays a role in reward pathways.[1] The raphe nuclei contain serotonergic neurons and project widely throughout the brain. The periaqueductal grey is thought to play a role in pain suppression.[2] The red nucleus is involved with movement and contains many connections with the cerebellum.[3] The medial longitudinal fasciculus lies anterior to the periaqueductal grey and plays a role in coordinating eye movements.
Pons
The pons connects the medulla oblongata inferiorly to the midbrain superiorly. The anterior portion of the pons is convex and can be easily seen as a visible distention when viewing the brainstem anteriorly. The surface of the anterior distention contains the basilar groove, which is where the basilar artery rests. The posterior pons is connected to the cerebellum by the middle cerebellar peduncles, which are the largest of the cerebellar peduncles. The posterior portion of the pons forms the superior portion of the floor of the fourth ventricle. A groove is formed inferiorly where the pons meets the medulla from which cranial nerves VI, VII, and VIII emerge medially to laterally. Cranial nerve V, the largest cranial nerve, exits from the superior anterior lateral pons. Important nuclei of the pons include the cranial nerve nuclei covered in the nerves section, the locus coeruleus, and pontine nuclei. The neurons of the locus coeruleus produce norepinephrine and have projections that spread widely throughout the CNS. The locus coeruleus is located in the posterior lateral pons at the lateral border of the periaqueductal grey and is involved in the reticular activating system. The locus coeruleus also suffers compromise in Alzheimer disease.[4][8] The pontine nuclei are a collection of pontine motor nuclei in the anterior pons that have many connections with the cerebellum via the middle cerebellar peduncle and assist with coordinating movement and help to modulate breathing.[5]
Medulla Oblongata
The most inferior portion of the midbrain is the medulla oblongata, which connects the pons to the spinal cord. It meets the spinal cord at the level of the foramen magnum. The anterior portion of the medulla oblongata contains the pyramids. The pyramids carry motor fibers from the precentral gyrus, or motor cortex, to the grey matter of the spinal cord, where they synapse and continue to the muscles of the body through the peripheral nervous system. The pyramids contain a decussation caudally in which the majority of the motor fibers contained cross to the contralateral side of the body. The fibers that decussate become the lateral corticospinal tract in the spinal cord, the fibers that do not decussate become the medial corticospinal tract in the anterior portion of the spinal cord. The pyramids lie on either side of the anterior median fissure, a midline groove that continues caudally along the anterior portion of the spinal cord. Lateral to the pyramids are the olivary bodies that are part of the olivocerebellar system. The hypoglossal nerve, cranial nerve XII, emerges from the anterior surface of the medulla from the sulcus between the olivary bodies and the pyramids. Posterior to the olivary bodies is the postolivary groove. Cranial nerves IX (glossopharyngeal), X (vagus), and XI (accessory) emerge from the postolivary groove in superior to inferior order. Posterior to the postolivary grooves are the inferior cerebellar peduncles that connect the medulla to the cerebellum. The posterior portion of the medulla oblongata inferiorly connects to the spinal cord. The posterior median sulcus is located in the midline on the posterior aspect of the inferior medulla and continues caudally along the posterior spinal cord. The posterior median sulcus in the posterior midline of the spinal cord is flanked by visible localized prominences called the gracile tubercles. On each side, the gracile tubercle contains the gracile nucleus. Similar bilateral localized prominences called the cuneate tubercles (containing the cuneate nucleus on each side) are just lateral to each of the gracile tubercles on the posterior aspect of the spinal cord. The gracile and cuneate tubercles carry second-order neurons of the dorsal column-medial lemniscus system. The gracile nucleus carries fibers for the lower extremities and trunk, and the cuneate nucleus carries fibers for the upper body above T6 except for the face and ears. The superior area of the posterior medulla oblongata forms a portion of the floor of the fourth ventricle.[6]
Major Brainstem Tracts
The Reticular Formation
The reticular formation is found in the anterior portion of the brainstem and is composed of multiple tracts that have a large number of connections. The reticular formation extends from the spinal cord through the brainstem to the diencephalon. It receives input from various tracts including spinothalamic tracts, spinoreticular tracts, the dorsal column-medial lemniscus pathway, visual pathways, auditory pathways, vestibular pathways, and cerebelloreticular pathways. The reticular formation sends efferent fibers to the thalamic nuclei, cerebellum, red nucleus, corpus striatum, substantia nigra, hypothalamus, and subthalamic nucleus.[6] The vast connections of the reticular formation allow it to modulate many different functions; some of these include movement coordination, autonomic regulation of blood pressure, heart rate, and respiratory rate, postural reflexes, neuro-vegetative reflexes, and taste.[6] It also plays a role in wakefulness and sleep.[7]
The Motor Tracts
Corticospinal Tracts
The majority of the upper motor neurons of the motor tracts originate in the precentral gyrus. The corticospinal fibers descend through the posterior limb of the internal capsule to the crus cerebri and then down the anterior pons to the pyramids of the medulla. At the pyramids, the majority of the corticospinal fibers decussate and descend the spinal cord as the lateral corticospinal tract and eventually continue to supply motor innervation to the limbs and digits. The majority of corticospinal fibers that do not cross over at the medullary pyramids become the medial corticospinal tracts, located anteriorly in the spinal cord, and provide innervation to the muscles of the trunk.[8]
Corticobulbar Tracts
The corticobulbar tracts descend through the genu of the internal capsule and down through a similar course as the corticospinal fibers; however, the corticobulbar fibers exit this course and synapse at the appropriate cranial nerve nuclei at their respective levels. The majority of corticospinal fibers decussate, while only some of the corticobulbar fibers decussate, as described in the nerves section of this article. The corticobulbar tracts also contain connections with many of the sensory nuclei of the brainstem.[6]
The Sensory Tracts
Spinothalamic Tract or Anterolateral System
The spinothalamic tract is responsible for conveying pain and temperature information from the body to the brain. Peripheral neurons carry sensory information to the posterior column of the spinal cord. After synapsing in the spinal cord, the axons ascend two to three levels before decussating. After decussating, the fibers ascend as the lateral and anterior spinothalamic tracts in the anterior and lateral portions of the spinal cord. When the tracts ascend through the medulla, they merge to form the spinothalamic tract and course along the lateral portion of the medulla. The tract continues up the lateral portion of the anterior pons and midbrain to the ventral posterior lateral thalamus, where the axons synapse and continue up through the posterior limb of the internal capsule to enter the post-central gyrus of the cortex.[9]
Dorsal Column-Medial Lemniscus
The dorsal column-medial lemniscus tract is responsible for carrying afferent proprioception, fine touch, two-point discrimination, and vibration to the cortex from the body. Peripheral neurons carry sensory information to the posterior column of the spinal cord and ascend in the posterior portions of the spinal cord as the gracile fasciculus and cuneate fasciculus. The neurons in these fasciculi will synapse of the gracile nucleus and cuneate nucleus at the level of the inferior medulla, respectively. The second order neurons will decussate at the level of the medulla and become the medial lemniscus. The medial lemniscus maintains a medial position within the brainstem as it ascends to the ventral posterior lateral thalamus. After synapsing in the thalamus, the fibers continue through the posterior limb of the internal capsule to the post-central gyrus of the cortex.[10]
Trigeminal Lemniscus and Spinotrigeminal Tract
Pain and temperature sensory input from the face enters the brainstem via cranial nerve V. The fibers that carry this information enter the brainstem and descend parallel to the spinal trigeminal nucleus before synapsing in it. Their descent forms the spinotrigeminal tract. After these fibers synapse, they decussate to the contralateral side and ascend as a part of the trigeminal lemniscus.[6]
The trigeminal lemniscus carries sensory axons from the second-order neurons of the principal sensory nucleus of the trigeminal nerve, which contain discriminative touch and oral cavity proprioception. These neurons do not descend before synapsing after entering the brainstem. Most of these fibers decussate to the contralateral side on their course to the ventral posterior medial thalamus and then proceed to the post-central gyrus of the cortex. The fibers of the trigeminal lemniscus ascend the pons and midbrain posterior to the medial lemniscus.
Lateral Lemniscus
The lateral lemniscus carries auditory information from the cochlear nuclei at the level of the inferior pons superiorly to the superior olivary complex, nuclei of the lateral lemniscus, inferior colliculi, and eventually to the medial geniculate body, which sends the auditory information to the temporal lobes of the cerebral cortex. Some of the fibers of the lateral lemniscus decussate while others do not. The lateral lemniscus travels up the posterior lateral portion of the pons and is important for sensory input to the brain.[11]
Embryology
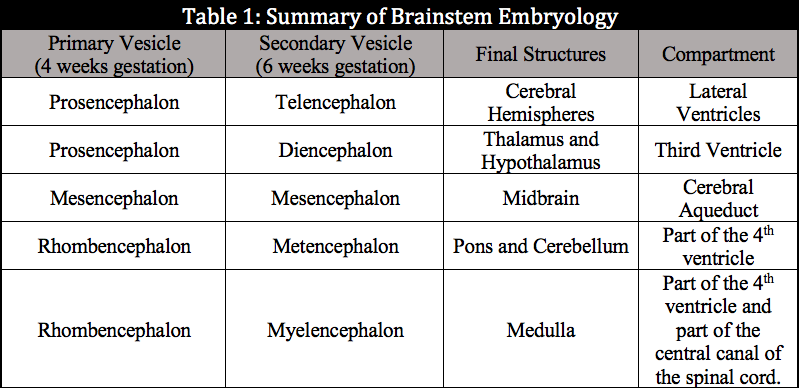
All of the CNS derives from ectoderm. It develops through the process of neurulation, which is induced by the notochord, which is of mesodermal origin. The notochord initially causes the formation of the neural groove and releases noggin and chordin to inhibit ectodermally derived bone morphogenic proteins and induce the creation of the neuroectoderm.[12] The neural fold is formed by about day 20 after conception and slowly folds over itself to form the neural tube. The tube closes both rostrally and causally, with the complete rostral closure occurring near day 24 post-conception and the caudal closure occurring near day 26 post-conception. Failure or other defects of the closure of the neural tube can result in various forms of spina bifida caudally and anencephaly, and craniorachischisis rostrally. At 4 weeks gestation, the neural tube will have developed three outpouchings that will give rise to the different components of the CNS. These outpouchings consist of the prosencephalon (forebrain), the mesencephalon (midbrain), and the rhombencephalon (hindbrain). By 6 weeks gestation, the prosencephalon will divide again into the telencephalon and diencephalon. The cerebral hemispheres and lateral ventricles arise from the telencephalon. The diencephalon will form the thalamus, hypothalamus, retina, and third ventricle. The mesencephalon will not divide any further and will give rise to the midbrain and cerebral aqueduct. At 6 weeks, the rhombencephalon will have divided further into the metencephalon and myelencephalon. The metencephalon will form the pons, cerebellum, and part of the fourth ventricle. The myelencephalon will produce the medulla and part of the fourth ventricle. The table below, Summary of Brainstem Embryology, contains this material in a simplified form.
Blood Supply and Lymphatics
The significant conduits of blood to the CNS are the internal carotid arteries and the vertebral arteries; they give rise to the many arterial branches that perfuse the CNS. The most inferior portion of the brainstem is the medulla oblongata. Caudally it receives the majority of its blood supply from the anterior spinal artery anterior-medially and the posterior spinal artery posterior laterally. Superiorly it receives the majority of its blood supply from the vertebral artery laterally, basilar artery branches anteriorly, and the posterior inferior cerebellar artery posteriorly.
Moving superiorly, the pons is the next brainstem structure encountered. Most of its blood supply comes from branches of the basilar artery. Superiorly, it is also perfused on its posterior lateral portion by branches of the superior cerebellar artery and branches of the anterior inferior cerebellar artery.
The next brainstem structure superior to the pons is the midbrain. It is perfused anterior medially by branches of the basilar artery, anterior laterally by branches of the posterior choroidal arteries and quadrigeminal artery originating from the posterior cerebral artery, and posteriorly by the quadrigeminal and superior cerebellar arteries.
The most superior brainstem structure, the diencephalon, is supplied anteriorly by branches of the anterior cerebral artery. Its posterior portions receive supply from branches of the posterior cerebral artery such as thalamogeniculate branches, thalamoperforating branches, and branches of the posterior communicating artery.
The anatomical lymphatic drainage of the central nervous system was described over 100 years ago, but until the last decade, the widely held consensus was that CNS lymphatics do not exist. The lymphatics of the CNS are small channels within the meninges and differ from the basic structure of the peripheral lymphatics. The CNS lymphatic system is still poorly understood but is thought to participate in immune cell transport, cerebrospinal fluid drainage, and interstitial fluid drainage. The lymphatics of the central nervous system continue to be a topic of investigation.[13]
Nerves
The entire brainstem is composed of neural tissue. Ten of the twelve cranial nerves also emerge from the brainstem.
Midbrain
The Oculomotor nerve (cranial nerve III) – Arises from the oculomotor sulcus on the medial portion of the crus cerebri. It is a motor nerve that receives inputs from two nuclei. The first nucleus is the oculomotor nucleus; it serves as its main motor nucleus and is in the anterior midline of the periaqueductal grey at the level of the superior colliculus. The second nucleus is the Edinger-Westphal nucleus, which provides parasympathetic motor inputs. The somatic motor fibers from the oculomotor nucleus provide innervation to all the extraocular muscles, with the exceptions of the superior oblique and lateral rectus muscles. The parasympathetic motor fibers of the Edinger-Westphal nucleus provide innervation to the ciliary muscles and constrictor pupillae after passing through the ciliary ganglion.
The Trochlear nerve (cranial nerve IV) – Exits from the posterior surface of the midbrain and is the only cranial nerve to exit posteriorly. It is a motor nerve with its nucleus located in the midline of the brainstem, also in the anterior portion of the periaqueductal grey, but inferior to the oculomotor nucleus. The nerve innervates the superior oblique ocular muscle, which is responsible for moving the eye downward and laterally. A unique feature of the trochlear nerve among the cranial nerves is that it is the only cranial nerve decussating peripherally. The nerve decussates at the superior medullary velum after leaving the brainstem, which causes cranial nerve nuclei deficits to appear as loss of function of the contralateral superior oblique muscle. Injuries that happen to the nerve distal to the decussation result in ipsilateral deficits to the superior oblique muscle.
Pons
The Trigeminal Nerve (cranial nerve V) – Arises from the superior anterior lateral pons as the largest cranial nerve. It contains both motor and sensory fibers. It arises as a smaller motor nerve and a larger sensory nerve. The sensory fibers provide innervation to the face and head. The motor fibers provide innervation to the muscles of mastication, mylohyoid, anterior belly of the digastric, tensor tympani, and tensor veli palatini. The trigeminal motor nucleus situates in the superior posterior lateral pons. The motor nucleus also receives corticobulbar fibers from both hemispheres as well as fibers from the reticular formation, medial longitudinal fasciculus, and red nucleus. The nerve has contributions from three sensory nuclei: The principal sensory nucleus of cranial nerve V, the mesencephalic nucleus, and the spinal trigeminal nucleus. The principal sensory nucleus of cranial nerve V lies directly lateral to the trigeminal motor nucleus and receives inputs from nerves that convey touch and pressure. The mesencephalic nucleus is located on the lateral aspect of the periaqueductal grey, anterior lateral to the fourth ventricle, and ascends to the height of the inferior colliculus. The mesencephalic nucleus conveys proprioceptive input from the teeth, hard palate, temporomandibular joint, and muscles of mastication. The spinal trigeminal nucleus is located in the inferior posterior lateral pons and extends inferiorly through the medulla into the superior spinal cord. The spinal trigeminal nucleus receives pain and temperature input for the sensory distribution of the trigeminal nerve.
The Abducens nerve (cranial nerve VI) – Is a motor nerve that emerges anteriorly and medially from the junction of the pons and medulla. The abducens nucleus is in the midline of the inferior tegmentum of the pons just ventral to the fourth ventricular floor. It provides innervation to the lateral rectus muscle, which is responsible for the abduction of the eye.
The facial nerve (cranial nerve VII) - Emerges from the junction of the pons and the medulla lateral to the abducens nerve at the cerebellopontine angle. It is both a motor and a sensory nerve and emerges as two separate roots; these include a medial motor root and a lateral sensory root. The facial motor nucleus situates in the anterior lateral inferior pons, just anterior and medial to the spinal trigeminal nucleus. The muscles of facial expression derive innervation from the motor nucleus of the facial nerve. The upper face receives corticobulbar fibers that partially decussate from both hemispheres, which allows sparing of deficits with lesions at the level of the cranial nerve nuclei while the lower muscles corticobulbar fibers fully decussate. The sensory nucleus is the upper portion of the solitary nucleus, which is located posterior and lateral to the facial nerve motor nucleus. It receives afferent fibers for taste from the anterior two-thirds of the tongue and sensation for the skin near the auricle of the ear. Its parasympathetic nucleus is the superior salivatory nucleus and is located laterally to the abducens nucleus but posterior to the facial motor nucleus. It innervates the submandibular and submental salivary glands.
The Vestibulocochlear nerve (cranial nerve VIII) – Arises from the brainstem directly lateral to the sensory root of the facial nerve. Cranial nerve VIII has two distinct portions, the vestibular, responsible for balance, and the cochlear, which is responsible for hearing. Cranial nerve VIII is purely a sensory nerve, and both of its portions course together until they reach their nuclei within the brainstem. The vestibular portion of the nerve provides input to the vestibular nuclei located along the lateral portion of the fourth ventricle in the inferior pons. The vestibular nuclei are composed of four different nuclei (superior, inferior, lateral, and medial.) These nuclei send tracts to three separate areas: the cerebellum via the vestibulocerebellar tract, the spinal cord through the vestibulospinal tract, and the nuclei of cranial nerves III, IV, and VI by the medial longitudinal fasciculus. The cochlear portion provides input to the dorsal and ventral cochlear nuclei. These nuclei are in the anterior lateral portion of the inferior pons. The posterior cochlear nucleus processes high-frequency sounds, while the anterior cochlear nucleus processes low-frequency sound. The anterior cochlear nucleus projects fibers to the ipsilateral superior olive and then to the lateral lemniscus. The posterior cochlear nucleus projects to the contralateral lateral lemniscus.
Medulla Oblongata
The glossopharyngeal nerve (cranial nerve IX) – Emerges from the postolivary groove and contains motor fibers, sensory fibers, and parasympathetic nerves. The nerve shares several cranial nerve nuclei with cranial nerve X (the vagus nerve), the nucleus ambiguus, and the nucleus solitarius. Cranial nerve IX also uses the inferior salivatory nucleus. The superior portion of the nucleus ambiguus is located posterior to the inferior olivary nucleus and contains second order motor cell nuclei for the glossopharyngeal nerve. The motor fibers from the superior nucleus ambiguus innervate the stylopharyngeus muscle. The cranial nerve IX parasympathetic cranial nerve nucleus is the inferior salivary nucleus; its postganglionic parasympathetic, visceral efferent fibers provide innervation to the parotid gland. The inferior salivary nucleus is located posterior to the nucleus ambiguus and receives afferent input from the hypothalamus, olfactory system, and nucleus solitarius. The last nuclei of the glossopharyngeal nerve are the nucleus solitarius. It is a sensory nucleus that receives taste from the posterior one-third of the tongue from the glossopharyngeal nerve. It also receives afferent impulses from the carotid sinus.
The Vagus nerve (cranial nerve X) – Emerges from the postolivary groove and contains motor fibers, sensory fibers, and parasympathetic fibers. The inferior portion of the nucleus ambiguus provides motor output to the muscles of the pharynx and larynx. The parasympathetic nucleus of the vagus nerve is the dorsal motor nucleus. The dorsal motor nucleus is located lateral to the hypoglossal nucleus and receives afferent fibers from the upper gastrointestinal tract, liver, pancreas, heart, and bronchi. The sensory nucleus for the vagus nerve is also the nucleus solitarius, similar to the glossopharyngeal nerve. It receives afferent inputs from the carotid sinus as well.
The accessory nerve (cranial nerve XI) – Arises from the medulla between the olive and inferior cerebellar peduncle and upper cervical spinal cord to C5. It forms from the combination of both cranial and spinal nerve roots. This nerve supplies both the trapezius and sternocleidomastoid muscles with motor innervation. Its efferent motor fibers arise from the nucleus ambiguus.
The hypoglossal nerve (cranial nerve XII) – Is a motor nerve and arises anteriorly from the medulla. Its nucleus sits in the midline of the brainstem anterior to the fourth ventricle. The hypoglossal nerve innervates the muscles of the tongue and the hyoglossus, genioglossus, and styloglossus muscles. It receives innervation via cortico-nuclear fibers from both hemispheres of the brain. However, the genioglossus muscle is innervated only by the contralateral side.[6]
Muscles
The brainstem is composed entirely of neural tissue and gets bathed in cerebrospinal fluid. The medulla oblongata joins the spinal cord at the level of the foramen magnum and is not in close contact with any muscles. The brainstem contains the cranial nerve nuclei for cranial nerves III-XII; therefore, the innervation of the muscles controlled by the previously mentioned cranial nerves are dependent on the brainstem.
Physiologic Variants
For supporting functions crucial to life and homeostasis like breathing, heart rate, sleep, and consciousness, the brainstem is of critical importance. The brainstem also is the most primitive portion of our brains. It is highly conserved with few differences in basic structure between vertebrates.[14] Because of its importance to life variants in brainstem anatomy or pathophysiology result in pathology with noticeable deficits. Examples of this include Arnold-Chiari malformations, with both types II and III involving the brainstem. Reports also exist of brainstem variants of hypertensive encephalopathy.[15] There are other rare cases of brainstem variants that result in deficits.
Surgical Considerations
Brainstem surgery may be necessary in cases of brainstem glioma and other tumors. These lesions can result in obstruction of cerebrospinal fluid (CSF) flow and obstructive hydrocephalus. Tumor size and location are considered in determining whether a patient is an operative candidate and in selecting the surgical approach.[16]
In cases of Arnold-Chiari malformations, a posterior fossa decompression surgery can be performed to make space for the brainstem and cerebellum. Brainstem access can be approached in many different ways depending on the desired exposure point. Some of these approaches include suboccipital, subtemporal, interhemispheric, transoral, and more. Any surgical procedure has inherent risk, and care must be taken during brainstem surgery not to damage the CNS and overlying structures while obtaining access.
Clinical Significance
An understanding of the anatomical location of brainstem structures and their blood supply is critical for localizing lesions on physical exam. Significant clinical problems can affect the brainstem such as stroke, malignancy, demyelinating processes, and many more. Important conditions that can affect the upper brainstem include but are not limited to thalamic pain syndrome from damage to the posterior lateral thalamus, hemiballismus from damage to the subthalamic nucleus, and changes in appetite, temperature regulation, blood pressure, growth and many more can suffer disruption by damage to the hypothalamus. Injury to or degeneration of dopaminergic neurons in substantia nigra results in Parkinson’s disease. Damage to the lower brainstem can result in many different syndromes that can be localized using deficits in cranial nerves and the tracts that traverse the brainstem, including but not limited to:
- Wallenberg stroke (spinothalamic tract, spinal trigeminal tract, hypothalamospinal tract, vestibular nuclei)
- Cerebellar tonsillar herniation (sudden respiratory and cardiac arrest due to compression of the medulla)
- Medial pontine syndrome (abducens nerve, corticospinal tract, medial lemniscus)
Another brainstem lesion that can occur is central pontine myelinolysis from the rapid correction of hyponatremia, which can result in seizures, ataxia, and disturbed consciousness.
Other Issues
Locked-in syndrome or pseudocoma is a rare neurologic condition that occurs when damage to the brainstem occurs, most commonly caused by ischemic or hemorrhagic stroke resulting in damage to the corticobulbar, corticopontine, and corticospinal tracts. This syndrome can result in patients with quadriplegia and dysarthria but who maintain consciousness. They may appear to be in a persistent vegetative state, but some patients may be able to communicate with eye movements or blinking and are aware of their surroundings but cannot communicate freely. Patients can live for years with this syndrome.[17]